|
|
|
|
|
Galleria Tassonomica
di
Natura Mediterraneo
|
|
|
| Autore |
 Discussione Discussione  |
|
Vitalfranz
Utente Senior


Città: Luxembourg
Regione: Luxembourg

2500 Messaggi
Flora e Fauna |
|
|
Hemerobius
Moderatore


Città: Alghero
Prov.: Sassari
Regione: Sardegna
4877 Messaggi
Flora e Fauna |
 Inserito il - 11 agosto 2008 : 20:51:57 Inserito il - 11 agosto 2008 : 20:51:57


|
| Messaggio originario di Vitalfranz:
...ma Roberto, ti volevo chiedere, tra specie vicariante e sottospecie che differenza c'è?
Secondo alcuni (l'ho visto citato altre volte) ci dovrebbe essere una stretta zona di sovrapposizione. Ma in questo modo sparirebbero tutte le sottospecie insulari...
Secondo me le specie dovrebbero essere ininterfeconde (o dare individui sterili), mentre le sottospecie dovrebbero essere feconde tra loro. Ma chi mai controlla questa cosa?
|
Ci sarebbe da scrivere un volume.
Sintetizzando molto, l'idea che mi sono fatto io è questa.
Due sottospecie hanno areali vicarianti ma nei punti di contatto esistono fasce più o meno larghe (fino a quanto larghe non si sa) con popolazioni intermedie. Naturalmente tra le due popolazioni che entrano in contatto vi deve essere scambio genico ma anche una discontinuità morfologica. In altre parole di qua una serie di caratteri che non si trovano di là, non una variazione "continua" dei caratteri (naturalmente esclusi gli individui intermedi). Questa situazione ha normalmente origini biogeografiche, due popolazioni di una specie rimangono isolate e poi rientrano in contatto. I due pool genici sono diversi, ma le due popolazioni non hanno meccanismi di isolamento riproduttivo, neanche in natura.
In due specie vicarianti c'è la medesima situazione, ma non c'è più possibilità di scambio genico perché le due popolazioni non sono in grado di riprodursi in natura.
In realtà la cosa non è così semplice perché dalla possibile riproduzione alla non-riproduzione ci sono tutta una serie di passaggi intermedi. Inoltre è assai comune che specie che in natura si ignorano nel 99,99% dei casi, messe in promiscuità in laboratorio (maschio di una e femmina dell'altra) non solo si accoppiano, ma producono anche prole fertile.
In questi casi la discrezionalità interpretativa del tassonomo è fondamentale, ma sempre deve essere giustificata. Non è giustificato invece istituire una sottospecie, anziché una specie, solo perché le differenze sono piccole. Non è il livello di differenziazione morfologica a rendere due popolazioni specie distinte o meno.
Per quanto riguarda la popolazioni insulari, il problema è insolubile. Se le capacità di dispersione della specie considerata ne impediscono la "evasione" dall'isola tanto vale, mia opinione personalissima, considerarle specie distinte. In questo caso la differenziazione morfologica rientra dalla finestra.
Comunque, come diceva Marcello La Greca, nell'istituire una sottospecie c'è una responsabilità molto maggiore che nell'istituire una vera specie.
Per ultimo la nomenclatura linneana è per sua natura "fissista" e mal si adatta ad una situazione "evoluzionistica". Ma, altra mia opinione personalissima, è ancora il miglior sistema che abbiamo. Qualcuno ci salvi dal BarCode.
Ciao Roberto 
verum stabile cetera fumus |
Modificato da - Hemerobius in data 11 agosto 2008 20:57:07 |
 |
|
|
gomphus
Moderatore

Città: Milano
Prov.: Milano
Regione: Lombardia
10487 Messaggi
Tutti i Forum |
Inserito il - 12 agosto 2008 : 09:49:44
|
caro roberto
vorrei aggiungere qualche considerazione alle tue, che condivido, sul concetto di sottospecie
pienamente d'accordo che per le popolazioni insulari, incluse le situazioni di "insularità" in aree continentali dovute ad es. alla localizzazione in ambienti relitti, il problema è insolubile, fra l'altro in moltissimi casi l'allevamento attualmente non è possibile; il fatto è che, nel processo di speciazione di due popolazioni con origine comune e in seguito riproduttivamente isolate, non esiste un momento in cui esse smettono di essere due semplici "popolazioni" per diventare due sottospecie, né uno in cui smettono di essere sottospecie e diventano specie a tutti gli effetti; si tratta di un percorso del quale ci troviamo in molti casi a osservare un singolo momento la cui esatta posizione resta imprecisata e probabilmente imprecisabile
allo stesso modo, immagino che non esista un momento in cui le due popolazioni divengono non più interfeconde: si andrà da un 10% di prole sterile a un 20, 30... fino al 100%, e lo stesso quando è l'accoppiamento stesso a non produrre prole, cioè non si ha nemmeno F1 (incidentalmente, ne conosco almeno un caso citato in letteratura riguardante quelle che tutti considerano due sottospecie), si comincerà con una piccola percentuale di accoppiamenti sterili che via via andrà aumentando
ad aggravare il problema, un fattore che pochi sembra tengano presente quando si ragiona di sottospecie e della loro validità, e che invece credo sia spesso incombente: ossia la possibile introduzione accidentale e successiva acclimatazione di una sottospecie in quello che fino a quel momento era l'areale di un altra, con possibile rimescolamento di caratteri e comparsa di forme intermedie o al limite di individui apparentemente tipici della sottospecie "sbagliata" (i caratteri distintivi possono anche essere specificati da geni dominanti), in aree tali da non potervi riconoscere alcun significato biogeografico
e se ad esempio nel caos che ruota intorno alla Megascolia maculata, che ho seguito su FNM senza ritenere di avere alcuna competenza per dire la mia, all'origine ci fossero state due sottospecie, chiamiamole "haemorrhoidalis" in Grecia e "flavifrons" in Italia, con successiva introduzione accidentale di "haemorrhoidalis" dalla Grecia all'epoca della colonizzazione della Magna Grecia, ad esempio come larve nei pani di terra con cui venivano trasportati gli alberi da frutta o di olivo da mettere a dimora nei futuri insediamenti?
aspetto i tuoi commenti
ciao
maurizio |
|
|
|
Hemerobius
Moderatore
Città: Alghero
Prov.: Sassari
Regione: Sardegna
4877 Messaggi
Flora e Fauna |
Inserito il - 12 agosto 2008 : 12:20:56
|
| Messaggio originario di Hemerobius:
In realtà la cosa non è così semplice perché dalla possibile riproduzione alla non-riproduzione ci sono tutta una serie di passaggi intermedi. |
| Messaggio originario di gomphus:
... il fatto è che, nel processo di speciazione di due popolazioni con origine comune e in seguito riproduttivamente isolate, non esiste un momento in cui esse smettono di essere due semplici "popolazioni" per diventare due sottospecie, né uno in cui smettono di essere sottospecie e diventano specie a tutti gli effetti; si tratta di un percorso del quale ci troviamo in molti casi a osservare un singolo momento la cui esatta posizione resta imprecisata e probabilmente imprecisabile.
allo stesso modo, immagino che non esista un momento in cui le due popolazioni divengono non più interfeconde: si andrà da un 10% di prole sterile a un 20, 30... fino al 100%, e lo stesso quando è l'accoppiamento stesso a non produrre prole, cioè non si ha nemmeno F1 (incidentalmente, ne conosco almeno un caso citato in letteratura riguardante quelle che tutti considerano due sottospecie), si comincerà con una piccola percentuale di accoppiamenti sterili che via via andrà aumentando. |
Qui diciamo assolutamente la medesima cosa (tu con maggiori particolari).
| Messaggio originario di gomphus:
ad aggravare il problema, un fattore che pochi sembra tengano presente quando si ragiona di sottospecie e della loro validità, e che invece credo sia spesso incombente: ossia la possibile introduzione accidentale e successiva acclimatazione di una sottospecie in quello che fino a quel momento era l'areale di un altra, con possibile rimescolamento di caratteri e comparsa di forme intermedie o al limite di individui apparentemente tipici della sottospecie "sbagliata" (i caratteri distintivi possono anche essere specificati da geni dominanti), in aree tali da non potervi riconoscere alcun significato biogeografico.
e se ad esempio nel caos che ruota intorno alla Megascolia maculata, che ho seguito su FNM senza ritenere di avere alcuna competenza per dire la mia, all'origine ci fossero state due sottospecie, chiamiamole "haemorrhoidalis" in Grecia e "flavifrons" in Italia, con successiva introduzione accidentale di "haemorrhoidalis" dalla Grecia all'epoca della colonizzazione della Magna Grecia, ad esempio come larve nei pani di terra con cui venivano trasportati gli alberi da frutta o di olivo da mettere a dimora nei futuri insediamenti?
|
Questo è un problema trascurato ma che ho ben presente. Tra l'altro le navi nell'antichità non viaggiavano mai vuote, se non avevano carico zavorravano la stiva con terra. Ti puoi immaginare quanta roba sia stata trasportata in questo modo. La localizzazione di alcuni Carabidi, Gasteropodi, Piante ed anche Tricladi, Collemboli, ecc. (cito gruppi su cui ho qualche dubbio) andrebbe letta anche in questa ottica.
La situazione è estremamente complessa. Però nel caso di un'introduzione accidentale di una sottospecie alloctona, visto che è altamente probabile sia avvenuta con un piccolo numero di esemplari, è altrettanto probabile che i geni tipici di questa vengano diluiti nella popolazione della sottospecie autoctona senza causare gravi problemi. Bisognerebbe altrimenti pensare che si creano "enclave" della sottospecie B nell'areale della sottospecie A. Perché? Mi vengono solo due risposte. Nella prima la sottospecie B "sbarca" in un territorio libero dalla sottospecie A (magari per motivi climatici) e crea una popolazione "indipendente" che solo successivamente entra in contatto con la sottospecie A inquinandola geneticamente in modo significativo. Ma più tirata per i capelli di così ...
La seconda è che siano due specie diverse!
Ciao Roberto
PS: non sono convintissimo di essere stato chiaro  ! !
verum stabile cetera fumus |
|
|
|
gomphus
Moderatore
Città: Milano
Prov.: Milano
Regione: Lombardia
10487 Messaggi
Tutti i Forum |
Inserito il - 12 agosto 2008 : 13:35:31
|
ciao roberto
sei stato chiarissimo!
vorrei solo aggiungere: nel primo caso il territorio colonizzato inizialmente dalla sottospecie B potrebbe essere libero, o quasi, dalla sottospecie A non per motivi climatici o comunque naturali, ma semplicemente perché è stato recentemente messo a soqquadro dall'insediamento umano ivi creato; così la sottospecie B può stabilirvi, come dire, una "testa di ponte" raggiungendo un numero non trascurabile di effettivi, irradiandosi successivamente tutt'intorno
se qualcuno dei caratteri distintivi di B è specificato da geni dominanti o anche incompletamente dominanti, per quanto diluito non scomparirà mai completamente, a meno che non sia controselezionato per qualche motivo, ad esempio espressione secondaria di una caratteristica metabolica o altro che nel territorio "nuovo" non è così vantaggiosa, oppure caratteristica cromatica, morfologica o biochimica (feromoni) insolita che interferisce con i meccanismi di riconoscimento fra i partner, ecc.
nel secondo... be', più o meno è dove volevo arrivare a proposito delle Megascolia, anche se esitavo a dirlo ad alta voce! suppongo che non solo nessuno (correggimi se ho sbagliato) abbia provato ad incrociare "haemorrhoidalis" e "flavifrons" e allevare la (eventuale) progenie per verificarne la fertilità, ma che ugualmente nessuno abbia fatto analisi di distanza genetica fra le due
in effetti, troppo spesso si rimane ancorati alla vecchia tassonomia morfologica per cui, se non vi sono differenze morfologiche rilevabili ma solo cromatiche, non possono essere due specie ma sono semplicemente due varietà (?)... e le sibling species allora? se esistono specie distinte le une dalle altre per caratteri biologici e non interfeconde, ma indistinguibili nell'aspetto, ugualmente possono esisterne che differiscono esteriormente solo per particolarità cromatiche
a questo proposito: a quanto ti risulta, in Italia meridionale e Sicilia si trovano anche forme intermedie, ad esempio con estensione variabile della pubescenza rossiccia, o si tratta di esemplari apparentemente ben caratterizzati di "haemorrhoidalis" occasionalmente rinvenuti insieme alle "flavifrons"? parlo di Italia meridionale e Sicilia perché gli esemplari francesi cui ho visto accennare, con solo l'estremità dell'addome rossiccia, potrebbero anche essere altra cosa, e qui si torna al discorso delle distanze genetiche fra popolazioni apparentemente ben diverse o al contrario apparentemente del tutto simili, distanze che quasi nessuno si preoccupa di verificare
ciao
maurizio |
|
|
|
Velvet ant
Moderatore

Città: Capaci
Prov.: Palermo
Regione: Sicilia
3768 Messaggi
Flora e Fauna |
Inserito il - 12 agosto 2008 : 13:40:22
|
Dato l'interesse generale assunto da questa discussione, ho ritenuto opportuno scinderla dal post da cui era partita ed inserirla, in evidenza, nel forum generale sugli insetti, in modo che abbia la massima visibilità e consenta una maggiore partecipazione da parte di tutti gli interessati.
Grazie a tutti per gli eccellenti interventi
P.S. - La discussione sulla Megascolia, dove pure appaiono alcune interessanti considerazioni sulle sottospecie e sulle variazioni intraspecifiche e a cui fa riferimento Maurizio (gomphus) nel pecedente intervento, è questa.
Anche questa discussione su Polyommatus daphnis pallidecolor mi pare si riallacci in qualche modo al tema trattato.
In questo caso, come già sottolineavo, ho l'impressione che, a parte quelle che sono le reali difficoltà di inquadramento e di definizione di questa categoria tassonomica, esista nei fatti un suo uso (e/o un abuso) non univoco e molto soggettivo, a seconda dei gruppi oggetto di studio (vedi quanto scritto in quella discussione).
Marcello


|
|
|
|
Hemerobius
Moderatore
Città: Alghero
Prov.: Sassari
Regione: Sardegna
4877 Messaggi
Flora e Fauna |
Inserito il - 12 agosto 2008 : 14:45:57
|
| Messaggio originario di Velvet ant:
Anche questa discussione su Polyommatus daphnis pallidecolor mi pare si riallacci in qualche modo al tema trattato.
In questo caso, come già sottolineavo, ho l'impressione che, a parte quelle che sono le reali difficoltà di inquadramento e di definizione di questa categoria tassonomica, esista nei fatti un suo uso (e/o un abuso) non univoco e molto soggettivo, a seconda dei gruppi oggetto di studio (vedi quanto scritto in quella discussione).
|
La discussione segnalata da Marcello mi era sfuggita. Per chi avrà la pazienza di leggerla (o per chi già la conosce) mi viene da dire che riterrei meno sbagliato (in mancanza di dati certi) considerare P. pallidecolor buona specie.
Questo esempio mi dà il destro per ribadire che il concetto di sottospecie è molto impegnativo ma con una sua base biologica. Sarebbe ora di smetterla di descrivere sottospecie solo sulla base di "piccole" differenze morfologiche. NON è il livello di differenziazione morfologica a fare di due taxa specie o sottospecie distinte.
I fattori biogeografici e lo studio della variabilità locale devono essere considerati in queste decisioni. Anche perché le analisi genetiche non sono così semplicemente attuabili e le prove di incrocio in laboratorio, se positive, danno quasi sempre informazioni slegate rispetto a ciò che avviene in natura.
Proprio per questo rubo tre righe a FNM per richiamare una indicazione bibliografica che avevo riportato nella discussione sulla Megascolia:
Marcello LA GRECA - L’uso delle categorie sistematiche sottogenere e sottospecie in tassonomia, alla luce della ricerca biogeografica. - Bollettino dell'Istituto di Entomologia "Guido Grandi" della Università degli Studi di Bologna 41 - 1987 p. 159-171.
Chi può se la procuri, ne vale la pena!
Ciao Roberto
verum stabile cetera fumus |
Modificato da - Velvet ant in data 12 agosto 2008 14:56:41 |
|
|
|
gomphus
Moderatore
Città: Milano
Prov.: Milano
Regione: Lombardia
10487 Messaggi
Tutti i Forum |
Inserito il - 12 agosto 2008 : 15:38:26
|
a roberto
perfettamente d'accordo, in questa situazione sarebbe meglio, in mancanza di dati precisi considerare P. pallidecolor come specie distinta.
in fin dei conti, in presenza di una popolazione ben isolata, con caratteri differenziali costanti rispetto a quelle italiane continentali, in mancanza di informazioni risolutive la trattazione come specie distinta non è particolarmente impegnativa, in quanto si può al limite considerarla un'ipotesi di lavoro piuttosto che una certezza acquisita; invece l'inquadramento come sottospecie, come giustamente sottolinei, è una scelta decisamente impegnativa che andrebbe adeguatamente motivata!
a marcello
concordo sul fatto che molti lepidotterologi hanno, come dire, la "sottospecie facile", però ti ricordo che alcuni nostri colleghi imenotterologi fanno anche di peggio! mi riferisco al fatto di accettare, anzi di dare per scontata, l'esistenza di "sottospecie ecologiche" (magari in casi in cui dell'ecologia e della biologia dei taxa coinvolti non si sa NULLA), e quindi di accettare come perfettamente logica l'idea di due o PIU' sottospecie della stessa specie contemporaneamente nella medesima località! (leggere per credere: W. Linsenmaier, 1959, Revision der Familie Chrysididae, Mitteilungen der Scweizerischen Entomologischen Gesellschaft 32 (1), p. 1). e non si tratta solo di idee del passato, perché alcune recentissime pubblicazioni sui crisidi riprendono gli stessi concetti...
ciao a tutti
maurizio |
|
|
|
Vitalfranz
Utente Senior
Città: Luxembourg
Regione: Luxembourg
2500 Messaggi
Flora e Fauna |
Inserito il - 13 agosto 2008 : 14:23:29
|
Sono molto contento di questa discussione, perché vedo che da posizioni un po' diverse, ma sostenute con cognizione di causa, si arriva piano piano a dipanare la matassa.
Riguardo alle specie insulari, sono solo in parte d'accodo con Roberto, perché bisognerebbe a questo punto distinguere tra isole "in allontanamento" per motivi geologici, e isole "vicine", ricollegate (o ricollegabili) durante la glaciazioni.
È noto che isole come l'Elba o il Giglio (o altre separate con fondali sotto i 100 m) erano collegate fino a 12000 anni fa da ponti di terra. Ponti che si sono formati e perduti più volte.
Studiando i cerambici del pino silvestre (lavoro ormai di oltre 20 pp. che prima o poi pubblicherò) mi son reso conto che le specie dei coleotteri sono invariate da almeno mezzo milione di anni e che perció isole come quelle citate prima NON POSSONO avere sottospecie particolari (tipo quelle descritte da Canzonieri).
Su altri libri ho letto che si stima che le specie di insetti non varino per almeno 7 milioni di anni e ciò corrisponde al fatto che la gran parte dei generi attuali si ritrova nel Baltico oltre 30 milioni da anni fa.
Il discorso si fa diverso per isole vulcaniche, come le Salomone, in cui l'introduzione di specie é avvenuta tramite uragani e l'isolamento é continuo (Vitali & Casadio, 2007).
In questo caso le specie tendono a differenziarsi e l'introduzione di nuovo materiale genico é dovuta solo agli uragani (e quindi al caso).
Perciò, quando si descrive un taxon (genere, specie o sottospecie), bisognerebbe fare molta attenzione se tutto quadra anche dal punto di vista geologico o paleologico, e non basarsi solo su differenze più o meno marcate o addirittura microscopiche.
Purtroppo - e s'è visto anche in qualche topic recente - si tende a stimare un entomologo solo in funzione del numero di taxa decritti.
È molto facile descrivere una specie nuova (se la si trova) e magari appiccicargli un nome senza badare troppo alle conseguenze...
Dimostrare veramente il rapporto tra due taxa dubbi in base ad allevamenti e prove d'incrocio porta via molto tempo e magari non fa metter il proprio nome in fondo ad un insetto. Forse per questo non lo fa quasi nessuno.
Tuttavia persone come Darwin e Fabre non hanno descritto nulla, ma il loro contributo alla storia dell'Entomologia (e dell'Umanità) è infinitamente superiore a quello di personaggi che hanno pubblicato lavori del tipo "60 nuove specie del Suriname"...
Francesco Vitali
|
|
|
|
gomphus
Moderatore
Città: Milano
Prov.: Milano
Regione: Lombardia
10487 Messaggi
Tutti i Forum |
Inserito il - 13 agosto 2008 : 16:02:37
|
ciao francesco
condivido quasi in pieno quanto dici sulle (sotto)specie insulari, ci sono solo un paio di punti sui quali non sono del tutto convinto:
1) riguardo alle specie delle isole vulcaniche o comunque separate da bracci di mare troppo ampi e profondi per essere emersi durante le glaciazioni (questa però magari è solo un'omissione), che la colonizzazione è avvenuta tramite uragani; aggiungerei: e, forse anche più spesso (soprattutto per quanto riguarda le isole mediterranee e la Macaronesia), su zattere di materiali vegetali finiti in mare in seguito a piene di fiumi o altro e trascinati dalle correnti, il che dovrebbe essere il meccanismo prevalente per insetti che quando hanno raggiunto le isole in questione erano già non volatori e magari micro-o anoftalmi
2) tu dici, a proposito dei cerambici, che secondo te "le specie dei coleotteri sono invariate da almeno mezzo milione di anni" e che secondo alcuni libri "si stima che le specie di insetti non variino per almeno 7 milioni di anni"; ritengo che non si possa generalizzare, in quanto esistono gruppi evolutivamente più stabili, più conservativi, come sembra siano i cerambici, e altri più plastici, come i carabidi. d'altra parte altri dati contraddicono quanto dici: secondo Machado (Monografía de los Carábidos de las Islas Canarias, 1992) a Fuerteventura si osserva una incipiente differenziazione morfologica tra le popolazioni di alcuni carabidi della penisola di Jandía e il resto dell'isola, ecologicamente separate da <22000 anni (non ancora abbastanza differenziate da meritare un nome a sé, ma è così che iniziano a formarsi le sottospecie e poi le specie), e lo stesso autore riporta che per due specie di Tenerife affini ma ben caratterizzate, Eutrichopus canariensis ed E. gonzalezi, localizzate sui massicci di Anaga e Teno (in passato rimasti a lungo isolati dall'attività vulcanica sulla parte principale dell'isola), Cobolli Sbordoni et al. 1987) hanno stimato su basi elettroforetiche una separazione risalente a 1,1 milioni di anni; mi riesce difficile accettare che se due popolazioni isolate da mezzo milione di anni sono ancora invariate e non differenziate, il successivo mezzo milione sia sufficiente a farne due buone specie
in ogni caso mi sembra che siamo lontani dagli "almeno 7 milioni di anni" durante i quali le specie di insetti non varierebbero, postulati dai libri che citi; potresti dirmi se i libri in questione citano dati concreti a sostegno di quanto affermano?
pienamente d'accordo invece nel lamentare che chi studia e poi descrive popolazioni localizzate su isole, e quindi per definizione (nella sua concezione) "isolate", troppo spesso non si pone minimamente il problema della profondità del mare fra l'isola e la costa, e quindi di "a quando" risale l'isolamento stesso; problema che personalmente sto considerando nello studio di alcuni carabidi nuovi delle Isole Ionie
concordo anche sul fatto che troppi considerano una misura del proprio successo come entomologi non la qualità complessiva della propria produzione, ma unicamente il numero di taxa descritti; ricordi il buon Maurice Pic, che si vantava di avere descritto 10000 specie, quando in effetti aveva semplicemente creato 10000 nomi (e neanche tutti diversi), lasciando ai posteri il compito di dipanare la matassa di cosa fosse valido e cosa no?
infine, per quanto riguarda le differenze morfologiche, se sono costanti e quindi ben caratterizzanti (senza entrare nel merito di "specie piuttosto che sottospecie"), sono d'accordo che il loro valore potrà essere anche questionabile, ma d'altra parte in molti, troppi casi attualmente non abbiamo molto altro, e quindi se hanno un riscontro in un quadro biogeografico, o altro, logicamente plausibile non possiamo non tenerne conto; vedi in proposito un precedente, e ottimo, intervento di roberto-hemerobius su questo post
"I fattori biogeografici e lo studio della variabilità locale devono essere considerati in queste decisioni. Anche perché le analisi genetiche non sono così semplicemente attuabili e le prove di incrocio in laboratorio, se positive, danno quasi sempre informazioni slegate rispetto a ciò che avviene in natura."
ciao
maurizio |
|
|
|
Hemerobius
Moderatore
Città: Alghero
Prov.: Sassari
Regione: Sardegna
4877 Messaggi
Flora e Fauna |
Inserito il - 13 agosto 2008 : 17:11:29
|
Forse si ripetono sempre le stesse cose, ma su questo argomento credo sia meglio essere un po' tediosi piuttosto che farsi fraintendere.
Dagli ultimi interventi vorrei estrapolare un paio di punti riguardo ai quali possono non essere stato chiaro.
Non ho mai escluso le differenze morfologiche dal novero dei caratteri che distinguono due taxa del livello specie (cioé specie o sottospecie). Anzi sono fortemente convinto che anche nelle specie gemelle più vicine, potendole separare a priori con altri metodi, è possibile trovare piccole e (quasi) costanti differenze morfologiche. Noi stiamo lavorando proprio a trovare queste differenze nel genere Chrysoperla (Neurotteri Crisopidi), uno di quegli intricatissimi gruppi con numerose specie recentissime (sono specie che possono avere una speciazione simpatrica per barriere riproduttive di tipo etologico, in questo caso i canti nuziali), e queste differenze le stiamo trovando.
Posso dire che rimango sempre molto perplesso (senza esprimere altre opinioni perché non conosco i gruppi) quando qualche collega coleotterologo mi dice: "Ho trovato il terzo esemplare noto di una specie rarissima, identica ad un'altra comunissima che si distingue solo dai genitali. Una faticaccia fare tutti quei preparati." Ma dove vive questa specie rara? Perché è rara? Qual'è il grado di differenziazione nei genitali? Possono esserci "mostruosità" dovute a, ad esempio, basse temperature che risultano costanti a parità di condizioni anomale (nei miei Rafidiotteri ne ho la convinzione)?
Detto questo, ribadisco anche che non è la "quantità" di differenziazione morfologica che può aiutarci nel descrivere una sottospecie anziché una specie. Tra due sottospecie le differenze ovviamente non possono essere grandi, ma tra due specie le differenze POSSONO essere anche molto piccole, eppure essere specie diverse.
Il discorso delle isole si ricollega a questo. Se io trovo una popolazione insulare con caratteri costanti che la differenziano da altre popolazioni continentali (o di altre isole) come mi devo comportare? La mia risposta è: discrezionalmente (e non descrizionalmente  ). Valuterò il grado di differenziazione morfologica, il loro grado di variabilità, le possibilità di contatto con le popolazioni vicine (continentali o di altre isole) attraverso le normali (o periodiche) capacità di dispersione della specie e mi domanderò: è necessario istituire un nuovo taxon per questa popolazione insulare? E se sì dovrà essere specie o sottospecie? ). Valuterò il grado di differenziazione morfologica, il loro grado di variabilità, le possibilità di contatto con le popolazioni vicine (continentali o di altre isole) attraverso le normali (o periodiche) capacità di dispersione della specie e mi domanderò: è necessario istituire un nuovo taxon per questa popolazione insulare? E se sì dovrà essere specie o sottospecie?
A mio parere la descrizione di piccole popolazioni insulari come sottospecie ha poco significato. Meglio essere coerenti e descrivere specie separate. Se queste popolazioni differiscono da quelle continentali (o di altre isole) per delle minuzie meglio sarebbe segnalare la cosa senza descrivere nulla.
Il ragionamento di Francesco sulle isole non mi pare in contraddizione con quanto sto dicendo. Le caratteristiche dell'isola sono uno degli elementi (principali) da valutare.
In tutto questo discorso va tenuto inoltre presente che la saltuaria introgressione di pochi geni da specie vicine sembra essere un meccanismo abbastanza usuale negli insetti per aumentare la proprio variabilità genetica (un po' come i batteri). Porto alla vostra attenzione un'interessante discussione sugli ibridi nei Lepidotteri (farfalle). Il Prof. Balletto dell'Università di Torino afferma: il tasso d'ibridazione stimato fra specie relativamente prossime è del 2-5%.
Francesco ha ragione che troppo spesso la descrizione di una nuova specie è obiettivo troppo ambito. Ma io ho come figura "esemplare" la mia maestra, la Prof. Maria Matilde Principi (ho appena postato una sua foto) che, nonostante sia unanimemente considerata una dei massimi Neurotterologi del secolo scorso, ha descritto in tutta la sua carriera una sola specie (buona naturalmente) e denominato (non descritto) un solo genere. E devo aggiungere scritto solo 100 lavori!!! ... e qui lasciamo perdere ...
Ciao Roberto
verum stabile cetera fumus |
|
|
|
Vitalfranz
Utente Senior
Città: Luxembourg
Regione: Luxembourg
2500 Messaggi
Flora e Fauna |
Inserito il - 13 agosto 2008 : 17:43:39
|
Ti rispondo con un po' di difficoltà, perché dopo i punto 1 e 2, non hai messi altri... ma spero di riuscire lo stesso.
1) Diffusione per tronchi.
No. Questa era un'ipotesi fatta da Gressitt, ma è di una leggenda metropolitana .
Se guardi le correnti oceaniche (ad esempio nelle Salomone) non collegano nessuna isola, ma passano attraversano le Salomone in maniera perpendicolare da nordest a sudovest, da zone senza alcuna isola (a meno che si voglia credere ad un'introduzione dal Sud America...)
Le Isole della Micronesia son poi soggette a correnti contrarie provenienti dal mare aperto.
La diffusione attraverso tronchi vale solo per bracci di mare ridottissimi, anche considerando il fatto che un tronco lasciato in mare non consente la sopravvivenza di larve per colpa del salino.
A breve uscirà un lavoro sul genere Olethrius, in cui tutto questo é spiegato molto bene, con tanto di cartine e direzioni delle correnti.
2) Età delle specie.
Tutte le specie che hanno raggiunto la Corsica e la Sardegna via Elba sono passate attraverso profondità di oltre 300 m. Questo tipo di profondità é stato probabilmente superato durante la regressione Siciliana di 500.000 anni fa (non prima, secondo alcuni anche 1 milione). Eppure le specie sono identiche a quelle del continente, e nessuno si sognerebbe di fare sottospecie o specie nuove (parlo dei cerambici).
Le specie del Campidano, di origine africana, sono più vecchie e si parla della regressione Messiniana che ha fatto ponte con Nord Africa (4-5 Milioni di anni fa). E sono specie poco dissimili da quelle nordafricane.
Il libro dei 7 milioni di anni l'ho letto nella libreria entomologica
di Washington e non l'ho fotocopiato perché non aveva parti sui cerambici... e putroppo non so dirti di più...
Tuttavia mi torna perché molti generi (Nothorhina, Paracorymbia, Strangalia, Pogonocherus) erano già presente allora, con specie quasi identiche alle nostre. Considerando che son passati almeno 30 milioni di anni e la velocità di speciazione degli autori che citi, dovremmo avere per il mondo un numero di specie di questo genere ben maggiore....
Dati concreti... la fauna di Willershausen (3,6-1,8 milioni da anni fa) ha rivelato la presenza di Ergates faber e Rosalia alpina. Sono state descritte altre specie, ma quelle che sto considerando (altro studio che uscirà) sono in realtá identiche a specie ora presenti in Asia o ancora in Europa.
Il problema é che molto spesso si tende a credere che un taxon fossile sia del tutto nuova, mentre magari si é solo spostato qualche migliaio di km più in lá...
In ogni caso son d'accordo con te che esistono gruppi più o meno stabili, e credo anche che una specie colonizzatrice di un'isola subisca un'accelerazione di speciazione.
Il resto son d'accordo con te... tranne una cosa: se "i dati di laboratorio danno informazioni slegate" sarebbe meglio capire perché ciò avviene...
Francesco Vitali
P.s. Scusa Roberto, ma abbiamo scritto (tanto) insieme e leggo ora il tuo post |
|
|
|
Vitalfranz
Utente Senior
Città: Luxembourg
Regione: Luxembourg
2500 Messaggi
Flora e Fauna |
Inserito il - 13 agosto 2008 : 17:54:43
|
Quoto in pieno quello che hai detto e mi associo alla tua stima verso la Prof. Principi. Fai bene a citarla e a ricordare quello che ha fatto (e come l'ha fatto!) perché é esemplare.
| Messaggio originario di Hemerobius:
Posso dire che rimango sempre molto perplesso (senza esprimere altre opinioni perché non conosco i gruppi) quando qualche collega coleotterologo mi dice: "Ho trovato il terzo esemplare noto di una specie rarissima, identica ad un'altra comunissima che si distingue solo dai genitali. Una faticaccia fare tutti quei preparati." Ma dove vive questa specie rara? Perché è rara? Qual'è il grado di differenziazione nei genitali? Possono esserci "mostruosità" dovute a, ad esempio, basse temperature che risultano costanti a parità di condizioni anomale (nei miei Rafidiotteri ne ho la convinzione)?
|
Ricordo un personaggio (non dico chi é) che venne al Museo per regalare la sua collezione di insetti minuscolissimi: erano tutti olotipi e paratipi nuovi, e non aveva alcuna altra specie che non fosse descritta da lui...
Francesco Vitali
|
|
|
|
gomphus
Moderatore
Città: Milano
Prov.: Milano
Regione: Lombardia
10487 Messaggi
Tutti i Forum |
Inserito il - 13 agosto 2008 : 20:30:49
|
caro francesco
grazie per le tue precisazioni, anche se devo farti notare che se dopo i punti 1) e 2) non ne ho messi altri, è perché avevo parlato di "un paio" di punti che non mi convincevano. comunque:
1) non ho inteso parlare di singoli tronchi entro cui dovrebbero essere contenute larve xilofaghe, ma di ammassi fluitati di detrito vegetale grossolano (ramaglie e simili), anche di grandi dimensioni, in cui possono essersi rifugiati coleotteri adulti, sicuramente più resistenti delle larve e in grado di sopravvivere per periodi abbastanza lunghi in queste condizioni, nonché abbastanza mobili da abbandonare queste zattere di fortuna appena queste approdano su una qualche terra emersa. ammetto tranquillamente di non sapere nulla della biogeografia delle isole del Pacifico, dato che non mi sono mai occupato di faune non paleartiche e non ho intenzione di cominciare adesso; tuttavia per l'area che più interessa il nostro forum, la colonizzazione su zattere è ammessa da Augusto Vigna Taglianti (anche in scambi di opinioni a livello personale, in questo caso non via forum) e da altri autori riportati ad es. nella monografia di Machado sui carabidi delle Canarie. quindi mi concederai che per lo meno la mia convinzione si basa su fonti che dovrebbero essere autorevoli, non è solo un'idea balzana che ho partorito in un momento di alienazione mentale
personalmente, per quel che vale la mia opinione di fronte a quelle di studiosi come te e roberto, cioè secondo me NULLA o giù di lì, per un minuscolo carabide endogeo microftalmo, come taluni trechini o pterostichini, o anoftalmo, come gli anillini, il meccanismo di colonizzazione su zattere (ripeto, parlo solo dell'area mediterranea e macaronesica) mi sembra più plausibile di quello che vedrebbe l'intervento di un uragano a risucchiare questi animaletti dal suolo dove si trovavano per depositarli a destinazione; confermo che per quanto riguarda i coleotteri mi occupo solo di carabidi, e quanto ai cerambici per lo più è già tanto quando arrivo a riconoscere i generi.
2) forse i dati che citi non sono così in contraddizione con quelli che ho trovato io, ma piuttosto confermerebbero l'esistenza di velocità di differenziazione molto diverse, ad esempio, tra cerambici e carabidi, o almeno quelli qui considerati.
sono convinto anch'io che una popolazione insulare o comunque di piccole dimensioni, per effetto anche di fenomeni di deriva genetica, tenda comunque a differenziarsi più rapidamente di quanto faccia la popolazione di origine; nel nostro caso, l'accelerazione in un gruppo molto stabile come sembrano essere i cerambici potrebbe essere comunque modesta, e in uno più plastico come i carabidi risultare molto più significativa.
quanto al fatto che "i dati di laboratorio danno informazioni slegate", informazione che ho preso da roberto perché di dati miei non ne ho (non dispongo nemmeno di un laboratorio adeguato qualora volessi raccoglierne per conto mio), sono assolutamente d'accordo che bisognerebbe capire perché ciò avviene; voglio solo dire che in attesa di capirlo, mi sembra che questi dati vadano almeno per il momento considerati congiuntamente a quelli morfologici per tentare una valutazione, non ritenuti in qualche modo "più importanti" degli altri in caso di discordanza; in fin dei conti, quando due di noi hanno opinioni discordanti su qualche argomento, non è così semplice stabilre chi ha ragione, no?
un'ultima cosa: nel corso di queste nostre discussioni, non è certo che voglio vedere riconosciute a tutti i costi le mie ragioni, voglio semplicemente sfruttare le competenze di specialisti di alto livello (non è una sviolinata) per cercare di capire qualcosa di un argomento che mi appassiona molto, e su cui le mie competenze mi sembra di capire che le "mie" competenze siano molto più scarse di quel che mi piacerebbe
grazie per la tua pazienza, anche futura...
ciao
maurizio
P.S. quando il tuo lavoro sull'identità delle specie fossili con quelle attuali sarà pubblicato, anche se non mi occupo di cerambici, mi piacerebbe riceverne o al limite acquistarne una copia. |
|
|
|
Hemerobius
Moderatore
Città: Alghero
Prov.: Sassari
Regione: Sardegna
4877 Messaggi
Flora e Fauna |
Inserito il - 13 agosto 2008 : 20:44:10
|
| Messaggio originario di Vitalfranz:
[in risposta a Maurizio]
Il resto son d'accordo con te... tranne una cosa: se "i dati di laboratorio danno informazioni slegate" sarebbe meglio capire perché ciò avviene...
Francesco Vitali
|
Beh, questo la avevo scritto io, e Maurizio mi ha citato, quindi sono io che devo chiarire.
La frase originale era:
"I fattori biogeografici e lo studio della variabilità locale devono essere considerati in queste decisioni. Anche perché le analisi genetiche non sono così semplicemente attuabili e le prove di incrocio in laboratorio, se positive, danno quasi sempre informazioni slegate rispetto a ciò che avviene in natura."
La spiegazione è semplice. Se prendete due specie diverse e le mettete in due contenitori, maschio di A e femmina di B nel primo, il contrario nel secondo, e non ottenete nulla potete affermare che le due specie non sono interfeconde (naturalmente dovrete avere anche il controllo, AxA e BxB, ed ottenere prole!).
Se invece ottenete prole anche dagli incroci AxB e BxA (od anche da uno solo di questi) e le F1 sono interfeconde potete solo affermare che non esistono barriere nell'accoppiamento e riproduttive post-zigotiche, ma NON POTETE affermare che le due specie si accoppiano anche in natura (esclusa una minima percentuale di ibridazione che negli insetti è pressocché fisiologica).
Non potete perché le barriere pre-zigotiche (o pre-accoppiamento) in laboratorio non funzionano come in campo, anzi spesso, nelle condizioni sperimentali date, non funzionano proprio.
Esiste una bellissima prova di colleghi giapponesi (ma è un esempio tra i tanti) che hanno provato ad incrociare Chrysoperla nipponensis con Chrysoperla carnea. Il successo negli incroci diminuiva all'aumentare delle dimensioni delle gabbie di allevamento! Gabbie più grandi rendevano più efficienti le barriere naturali, basate in questo caso su canti nuziali specifici.
Quindi molto spesso il gioco non vale la candela, nel senso che la fatica e le risorse necessarie per queste prove (quando possibili, cioé quando siete in grado di riprodurre AxA e BxB, il che non è sempre detto) non forniscono risultati proporzionali allo sforzo compiuto.
Ciao Roberto
PS: vedo che, mentre rispondevo io, rispondeva anche Maurizio (è una vera corsa contro il tempo ), ma credo che il messaggio sia comunque comprensibile.
verum stabile cetera fumus |
|
|
|
gomphus
Moderatore
Città: Milano
Prov.: Milano
Regione: Lombardia
10487 Messaggi
Tutti i Forum |
Inserito il - 13 agosto 2008 : 21:20:33
|
caro roberto
anch'io sottoscrivo in pieno quanto hai detto, in particolare riguardo al fatto che le differenze fra due specie, se costanti, possono essere anche "piccole"; solo vorrei confermarti che per quanto mi riguarda mi sembra che tu sia sempre stato chiarissimo, e non credo che ci siamo fraintesi.
a proposito del livello di differenziazione di due specie, c'era una parte del ragionamento di francesco che non mi convinceva: "bisognerebbe fare molta attenzione... e non basarsi solo su differenze più o meno marcate". in pratica mi era sembrato di capire in questa ottica differenze poco marcate avessero a priori scarso valore, e questo mi era sembrato in contrasto con il tuo punto di vista, che ripeto condivido in pieno, che se le differenze sono costanti non importa se sono piccole. a questo punto credo però che si sia trattato solo di un equivoco da parte mia.
un'altra cosa: quello che ho appena scritto a francesco, "nel corso di queste nostre discussioni, non è certo che voglio vedere riconosciute a tutti i costi le mie ragioni, voglio semplicemente sfruttare le competenze di specialisti di alto livello (non è una sviolinata) per cercare di capire qualcosa di un argomento che mi appassiona molto, e su cui mi sembra di capire che le "mie" competenze siano molto più scarse di quel che mi piacerebbe" vale anche per te.
piuttosto, mi interessa moltissimo quello che hai citato, circa le possibili limitate introgressioni genetiche dovute a saltuari accoppiamenti ibridi; ci sono un paio di casi negli odonati (non ti annoio con i dettagli, a meno che non ti interessi) in cui le distanze genetiche forniscono risultati in evidente contrasto con quello che risulterebbe in base alla sistematica morfologica. i meccanismi che hai citato potrebbero essere una spiegazione.
anche il discorso su possibili anomalie a comparsa costante, in risposta a condizioni ambientali particolari, come possibile origine di specie rare, mi sembra molto interessante e da approfondire. un meccanismo simile per la verità era già stato ipotizzato riguardo a due specie (?) di Anax.
ciao e... grazie anche a te per la pazienza
maurizio
P.S. ti ho mandato stamattina un email sull'indirizzo di posta del CNR, l'hai trovato?
|
|
|
|
Vitalfranz
Utente Senior
Città: Luxembourg
Regione: Luxembourg
2500 Messaggi
Flora e Fauna |
Inserito il - 13 agosto 2008 : 21:22:10
|
Ringrazio entrambi per le preziose informazioni e le occasioni di riflessione che mi state offrendo.
Desidero anche ringraziarvi per il livello della discussione... chi qualche volta si é lamentato che determiniamo solo Oedemera nobilis  , potrà stavolta essere soddisfatto (spero , potrà stavolta essere soddisfatto (spero  ). ).
Il fatto che ci sovrapponiamo nelle risposte é un bellissimo segno.
Per Maurizio: il mio discorso sulle correnti valeva solo per le isole del Pacifico, (che mi sono studiato) per il resto (e soprattutto per la diffusione di insetti atteri, come i Carabidi, e nel Mediterraneo), il discorso tuo e di Vigna Taglianti é perfettamente plausibile.
Per citarti: non si può generalizzare mai.
Soprattutto sulla Natura: le regole sono talmente varie e complesse e il Mondo così grande che non ci si può stupire di nulla.
Per Roberto: ti ringrazio della notizia di questi esperimenti giapponesi.
Mi sembra che in condizione di cattività "saltino un po'" certe regole.
Sapevo di animali superiori (cani) che passano da eterosessuali a omosessuali in cattività. Forse il discorso va un po' oltre e me ne scuso, ma credo che sia una cosa un po' analoga.
P.s. Per l'articolo, mi farò onore di inviarti(vi) una copia appena pubblicato.
Se ti(vi) interessano altri articoli sui fossili, posso inviarti(vi) i Pdf anche subito. Le copie cartacee purtroppo ora no: qui non ho portato (tra le altre cose, di cui sa ormai tutta Italia....) neanche loro...
Francesco Vitali
P.p.s. Maurizio, ci siamo accavallati un'altra volta! |
|
|
|
Hemerobius
Moderatore
Città: Alghero
Prov.: Sassari
Regione: Sardegna
4877 Messaggi
Flora e Fauna |
Inserito il - 13 agosto 2008 : 22:45:31
|
| Messaggio originario di gomphus:
piuttosto, mi interessa moltissimo quello che hai citato, circa le possibili limitate introgressioni genetiche dovute a saltuari accoppiamenti ibridi; ci sono un paio di casi negli odonati (non ti annoio con i dettagli, a meno che non ti interessi) in cui le distanze genetiche forniscono risultati in evidente contrasto con quello che risulterebbe in base alla sistematica morfologica. i meccanismi che hai citato potrebbero essere una spiegazione.
|
Lo studio delle ibridazioni naturali è campo, a quanto ne so, quasi esclusivo dei lepidotterologi. Le mie informazioni sono di seconda mano, bisognerebbe sentire Balletto a Torino, o Sbordoni a Roma, e comunque un po' di letteratura "genetica". Io ho sottomano qualcosa che riguarda il genere Chrysoperla, ma poca cosa.
Attento che le distanze genetiche sono misure relative, non assolute. Se ci sono differenze contrastanti con la morfologia bisogna verificare cosa non va ... ed a volte è il "campionamento" dei geni (o del numero delle specie, che in ultima analisi è la stessa cosa) su cui si calcola la distanza ad essere carente.
Nei Neurotteri abbiamo fortunatamente la studiosa di punta dei rapporti filogenetici basati sulle distanze geniche che è stata morfologa. E dopo aver rivoluzionato qualcosa (non molto per il vero), ha appena pubblicato una sintesi in cui riaccetta tutte le "vecchie" concezioni alla luce dei "nuovi dati" ().
| Messaggio originario di gomphus:
anche il discorso su possibili anomalie a comparsa costante, in risposta a condizioni ambientali particolari, come possibile origine di specie rare, mi sembra molto interessante e da approfondire. un meccanismo simile per la verità era già stato ipotizzato riguardo a due specie (?) di Anax.
|
Io lo ritengo possibile in insetti piuttosto primitivi. Fra quelli che studio, me ne sono convinto nei Rafidiotteri, ma sono altrettanto convinto che sarebbe impossibile nei molto più evoluti Neurotteri (s.str. = Planipenni).
Mi sembra di aver visto recentemente qualcosa sul bollettino della SEI riguardante i Plecotteri.
Ciao Roberto
verum stabile cetera fumus |
|
|
|
gomphus
Moderatore
Città: Milano
Prov.: Milano
Regione: Lombardia
10487 Messaggi
Tutti i Forum |
Inserito il - 13 agosto 2008 : 23:44:16
|
ciao roberto
in effetti anch'io avevo pensato, nel primo caso (genere Chalcolestes in Italia), che qualcosa era andato storto nella scelta delle sequenze da paragonare o qualcosa del genere (scusa il modo di esprimermi, ma è un argomento che veramente conosco solo a grandi(ssime) linee); nel secondo caso però (gen. Platycnemis del bacino mediterraneo) è più difficile... per spiegare il fatto che le specie di una data area somigliano geneticamente (DNA mitocondriale) più a quelle morfologicamente dissimili e simpatriche che non a quelle apparentemente affini ma allopatriche, dato che i mitocondri come origine sono batteri-o-qualcosa-del-genere divenuti simbionti obbligati, cosa ne pensi dell'idea, che mi era venuta in mente ma che ho sentito recentemente avanzare anche da gente molto più preparata di me, di una possibile trasmissione "infettiva" delle sequenze mitocondriali tra specie diverse conviventi?
e con questo per stasera chiudo, a domani
maurizio |
|
|
|
Acipenser
Utente Senior

Città: Francoforte sul Meno
Prov.: Estero
Regione: Germany
1668 Messaggi
Tutti i Forum |
Inserito il - 14 agosto 2008 : 02:24:40
|
| Messaggio originario di gomphus:
ciao roberto
in effetti anch'io avevo pensato, nel primo caso (genere Chalcolestes in Italia), che qualcosa era andato storto nella scelta delle sequenze da paragonare o qualcosa del genere (scusa il modo di esprimermi, ma è un argomento che veramente conosco solo a grandi(ssime) linee); nel secondo caso però (gen. Platycnemis del bacino mediterraneo) è più difficile... per spiegare il fatto che le specie di una data area somigliano geneticamente (DNA mitocondriale) più a quelle morfologicamente dissimili e simpatriche che non a quelle apparentemente affini ma allopatriche, dato che i mitocondri come origine sono batteri-o-qualcosa-del-genere divenuti simbionti obbligati, cosa ne pensi dell'idea, che mi era venuta in mente ma che ho sentito recentemente avanzare anche da gente molto più preparata di me, di una possibile trasmissione "infettiva" delle sequenze mitocondriali tra specie diverse conviventi?
e con questo per stasera chiudo, a domani
maurizio
|
Ciao, ti rispondo solo sul lato molecolare. Trasmissioni orizzontali di materiale genetico non sono da escludersi nemmeno negli organismi superiori. Parlando di questo argomento si entra certamente nella biologia "profonda", si fanno cioè osservazioni su ciò che davvero può essere definito sistema biologico.
In effetti gli organismi fanno parte di un insieme più grande di "enti" che sono in grado di riprodurre sé stessi sfruttando ciò che trovano nell'ambiente, e mi spiego meglio: se consideriamo un batterio come parte dell'ambiente (e così è in ultima analisi) un plasmide che si replica utilizzando metaboliti e sistemi enzimatici del batterio altro non fa che comportarsi come un "vivente".
Di seguito vi propongo una classificazione (un documento da me prodotto per questo topic) dei viventi che forse potrà sembrare un po' atipica, ma che ritengo opportuno chiamare in causa per la piega che ha preso la discussione:
Immagine:
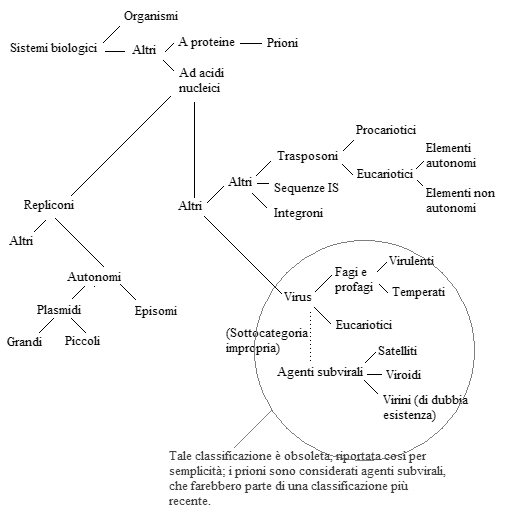
83,38 KB
Tautò tèni zon kài
tethnekós kai egregoròs
kai kathèudon kai nèon kai
gheraiòn tade gàr
metapésonta ekéina ésti
kakèina pàlin táuta.
Eraclito, Frammenti, 88; Di passaggio, Franco Battiato. |
|
|
|
Hemerobius
Moderatore
Città: Alghero
Prov.: Sassari
Regione: Sardegna
4877 Messaggi
Flora e Fauna |
Inserito il - 14 agosto 2008 : 07:13:57
|
| Messaggio originario di Acipenser:
| Messaggio originario di gomphus:
... per spiegare il fatto che le specie di una data area somigliano geneticamente (DNA mitocondriale) più a quelle morfologicamente dissimili e simpatriche che non a quelle apparentemente affini ma allopatriche, dato che i mitocondri come origine sono batteri-o-qualcosa-del-genere divenuti simbionti obbligati, cosa ne pensi dell'idea, che mi era venuta in mente ma che ho sentito recentemente avanzare anche da gente molto più preparata di me, di una possibile trasmissione "infettiva" delle sequenze mitocondriali tra specie diverse conviventi? ...
|
Ciao, ti rispondo solo sul lato molecolare. Trasmissioni orizzontali di materiale genetico non sono da escludersi nemmeno negli organismi superiori. Parlando di questo argomento si entra certamente nella biologia "profonda", si fanno cioè osservazioni su ciò che davvero può essere definito sistema biologico.
...
|
Grazie prima di tutto ad Acipenser per l'intervento graditissimo che porta un importante contributo di competenze, diverse da quelle dei (finora) principali partecipanti a questa discussione.
Spero che anche altri intervengano, magari con domande e dubbi o, se inesperti, con richieste di spiegazioni. Personalmente ne sarei ben felice perché le domande spesso insegnano più delle risposte.
L'ipotesi di Maurizio non mi sembra campata in aria anche se non ho le competenze genetiche (e le conoscenze particolari su questo "caso") per giudicare compiutamente. Quello che voglio sottolineare è che il dato genetico non è "verità rilevata", ma un elemento di conoscenza aggiuntivo (importantissimo) rispetto a tutti gli altri (morfologici, ecologici, biogeografici, &c). Spesso i dati genetici sono "campionati" bene ma in quantità insufficiente. Talvolta sono proprio campionati male!
Ciao Roberto
verum stabile cetera fumus |
|
|
|
Discussione |
|
|
|
 Natura Mediterraneo Natura Mediterraneo |
© 2003-2024 Natura Mediterraneo |
|
|
Leps.it | Herp.it | Lynkos.net
|

 Forum
|
Registrati
|
Msg attivi
|
Msg Recenti
|
Msg Pvt
|
Utenti
|
Galleria |
Map |
Forum
|
Registrati
|
Msg attivi
|
Msg Recenti
|
Msg Pvt
|
Utenti
|
Galleria |
Map |
